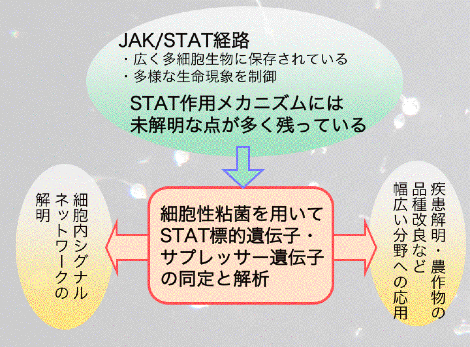
転写因子STATとは?
STATとはSignal Transducers and Activators of Transcriptionという名前の転写因子です。この転写因子はもともとインターフェロンやインターロイキンなどのサイトカインに応答する転写制御因子として哺乳類で見つかってきたもので、免疫応答に関与することが分かっていました。その後、さまざまな物質(例えば増殖因子やエリスロポイエチンなど)もリガンドとして作用することが分かってきました。STATは哺乳類のような高等な生き物にしか存在しないと当初は考えられていましたが、その後ショウジョウバエでも存在することが分かり、さらには細胞性粘菌にも存在することが分かり、今では真核多細胞生物に広く存在することが知られています。最近では、植物にも同様のアミノ酸配列を有するタンパク質をコードする遺伝子があることが分かってきました。しかしながら、全ゲノム配列が解読されている酵母にはSTATと思われるものは存在せず、多細胞生物に特異的なものと考えられています。
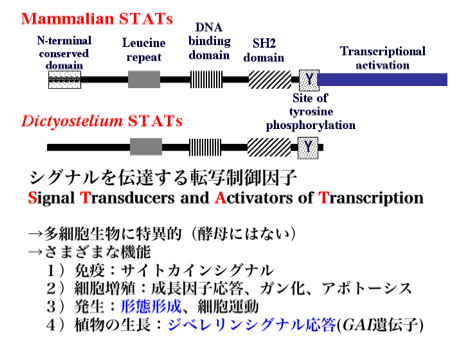
上図はSTATタンパク質の一次構造を模式的に示したもので、ちょっと古いものです。特徴的な構造はC末端側にSH2ドメイン(Src homology-2 domain)がそのすぐC末端側にはリン酸化されうるチロシン残基が存在し、両者ともに2量体形成に関わっています。N末端側は多量体形成に関わっているといわれ、中央にDNA結合ドメインが存在します。STATは特異的なDNAの塩基配列に結合し、転写を促進あるいは抑制します。細胞性粘菌のSTATには哺乳類などのSTATにみられるC末端の転写活性化ドメインに相当するものが見られません。しかしながら、遺伝子によっては明らかにSTATによって転写が活性化されるものが知られており、どのようなメカニズムによるのか未だに分かっていません。
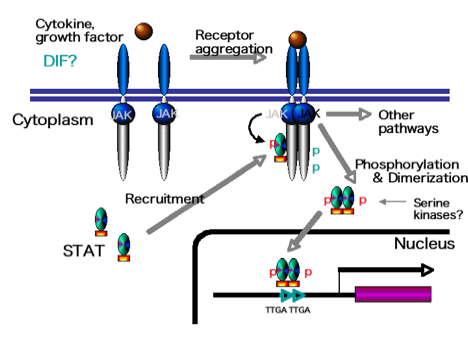
STATはJAK-STAT経路(下図)と呼ばれるシグナル伝達経路の最後に位置しています。サイトカインや増殖因子、細胞性粘菌の場合はcAMPやDIF-1(Differentiation inducing
factor-1)というような低分子の物質やタンパク質がリガンドとなって細胞表面の受容体に結合します。受容体が結合すると受容体分子どうしが寄り添って互いをリン酸化します。そうすると、細胞質で不活性な状態で存在していたSTATが受容体に引き寄せられ、受容体に付随しているJanusキナーゼ(JAK)によってSTATのSH2のすぐC末端にあるチロシンがリン酸化されます。リン酸化されたSTATは活性化され2量体を形成し、核へ移行し標的のDNA配列に結合し転写を調節します。この経路は、関わる分子に多少の違いは見られますが細胞性粘菌からヒトまで保存されています。
このように、細胞性粘菌からヒトまで関わる分子が保存されているということは、細胞性粘菌で調べたことが(いつもそうとは限りませんが)ヒトにおいても同じである可能性があることを示しています。STATはそれ自体がガン遺伝子としてはたらくなど非常に重要な生命現象を多く司っており、また、未解明な現象も数多く残されています。我々の研究室では細胞性粘菌でしか出来ないような大規模スクリーニングの手法をもちいて、進化的な関係も考慮に入れてSTATの機能を解析しています。