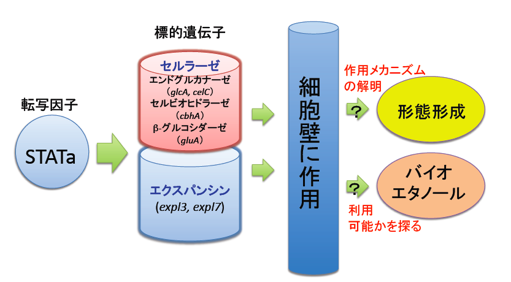
多細胞の真核微生物である細胞性粘菌はセルロース合成酵素を有し、セルロースが多細胞体の形態形成(体の形を作ること)において重要な働きをしています。また、細胞性粘菌(一般に粘菌類)はセルロース•バイオエタノールの産生において注目されているセルロース分解酵素(セルラーゼ)を多数有していて(Eichinger, et al., 2005)、多細胞体でセルロースが構築される時期と同時期に発現していることも知られています。既に、我々の研究室では幾つかのセルラーゼ遺伝子と植物細胞壁の弛緩作用を持つエクスパンシン遺伝子が転写因子STATaの標的遺伝子になっていること[テーマ2]や実際に1つのエクスパンシンが形態形成を制御していることを見いだしています(Ogasawara
et al., 2009)。このテーマは、これらのセルラーゼ遺伝子やエクスパンシン遺伝子の働きをより詳細に解析し、形態形成にどのようにして働いているのかを解明することを目的とします。また、ついでですが、セルロース•バイオエタノール産生における粘菌類利用の可能性についても検討します。
(1)植物や真菌(カビやキノコ、酵母菌)、およびバクテリア(細菌)の細胞は外側には丈夫な細胞壁が発達しています。細胞壁は、単にこれらの生物の細胞形態を決定して、物理的な強さを与えているだけでなく、細胞膜や表層細胞骨格(細胞の骨組み)と密接に相互作用して細胞内活動の制御に積極的に関わっていることが徐々に明らかになってきています(Mutwil, et al., 2008; Hématy and Höfte,
2008)。しかしながら、酵母や真菌類、一部の植物を除いて、細胞壁の詳しい生理機能は依然として不明の部分が多く残っています。その原因は、細胞壁の主要な構成成分であるセルロース(地球上に最も多く存在するバイオマス)やそれを分解するセルラーゼが、生物の種によって構造や機能がさまざまで統一的な理解が不十分なためで、多くの生物種からの情報を集めることと簡単にセルラーゼの生体における機能を調べることが出来るいわゆるバイオアッセイ系の開発が必要と思われます。
一方、真核多細胞微生物である細胞性粘菌も子実体形成過程においてセルロースからなる細胞壁を構築し、セルロースの合成が正常な形態形成に不可欠であることが示されています(Blanton et al., 2000)。我々は、子実体形成がどのような分子で制御されているのかを調べる過程で、STATaという転写因子が中心的な働きをしていることを明らかにしました(Kawata et al., 1997)。さらに、STATaの標的遺伝子を探索した結果、4つのセルラーゼ遺伝子と2つのエクスパンシン遺伝子がその中に含まれることが分かりました[テーマ2]。これらは、いずれもセルロースに働きかけるタンパク質をコードしていることから、これらの機能を調べれば、セルロースによる形態形成メカニズムの一端が見えてくると考え研究を始めています。
(2)バイオエタノールというものが石油の代わりのエネルギーとして注目されていますが、バイオエタノールを作るための原材料の多くはトウモロコシなどが作るでんぷんです。これらは食料でもあるため、食料価格が高騰するという欠点を伴います。そのため、現在世界中で食料と競合しない植物バイオマスを利用したバイオエタノールの開発が研究されています(Rubin, 2008)。バイオマスとして最も注目されているものが地球上に最も多く存在するセルロースです。変形菌(真性粘菌)や細胞性粘菌はもともと森林中に存在し、バクテリアなどを食すことから、ソフトセルロースを分解する能力を持つようです。廃材などの木質のセルロースを分解出来るかまでは分かりませんが、直接にセルロースを餌とし粘菌類を培養してセルロースを分解出来ると、セルロースバイオマスに対して余分な前処理が不要となり効率よくバイオエタノールを得ることが可能になると期待されます。ここでは、我々の研究室で解析している遺伝子を改変し、組み替え体を作製することでセルロースの分解能が変化するかどうかを検証しようとしています。
解析結果:
➡2011年度のまでの展開はこちら
<参考文献>
1. Eichinger, L., et al. (2005)
Nature 435:43-57.
2. Mutwil, M., et al. (2008) Curr.
Opin. Plant Biol. 11:252-257.
3. Hématy,
K. & Höfte, H. (2008) Curr. Opin. Plant Biol. 11:321-328.
4. Blanton, R. L., et al.,
(2000) PNAS 97:2391-2396.
5. Kawata, T., et al.
(1997) Cell 89:909-916.
6. Rubin, E.
M. (2008) Nature 454:841-845.
STATと細胞壁の機能に関して:
転写因子STATと相同なタンパク質は、植物においては植物ホルモンの1つであるジベレリンによって制御を受けることが知られています。また、細胞壁の弛緩作用を持つエクスパンシンもジベレリンによって制御を受けることが知られています。同じようなシグナル伝達経路が細胞性粘菌にも存在することは、細胞壁の新機能の発見やジベレリンによる植物の生長制御のメカニズムに関しての進化や比較生化学に関して有意義な情報が得られるものと予想されます。特に、細胞壁の主成分であるセルロースに関しては、セルラーゼ遺伝子の過剰発現などにより、形態形成におけるセルラーゼやセルロースの役割が明らかになると期待されます。さらに、セルラーゼによって細胞壁の微細な固さの調節が行われて、これが植物における茎頂分裂組織(apical meristem) の未分化状態を維持している可能性が指摘されています。植物の茎頂分裂組織はちょうど動物の幹細胞と同じような働きをしていると考えられ、どのようなメカニズムで未分化状態を維持しているのかという疑問の解明に役立つと思われます。
セルロースバイオマスの利用に関して:
バイオエタノールの生産系の代替え原料としてセルロースが最も注目されています。もともと森林中に存在し、セルロースを分解する能力を持つ粘菌類は、直接にセルロース原料に付着し分解する点でもっと注目されるべきと思います。本研究によって前処理の不要な効率の良いセルロース分解系の可能性が示されれば(そんなに簡単なことではないですが)、産業的に多大な貢献があると期待されます。